Чем полезна избирательность в отношении ориентации? Ответ все тот же: она экономит ресурсы благодаря уменьшению объема информации, передаваемой на следующие этапы зрительной обработки (в данном случае в высшие корковые отделы). Важно отметить, что это делается с сохранением значимой информации, необходимой для идентификации объекта. Самые важные визуальные стимулы – это объекты, а объекты определяются своими краями. Во многих случаях мозгу достаточно знать об ориентации краев – и ни о чем больше, – чтобы догадаться, какой перед ним объект.
На правом рисунке на следующей странице показаны только края – контуры собаки, – которые регистрируются простыми клетками первичной коры. Несмотря на то, что в контурном рисунке частично теряется богатство исходного изображения, собака остается легкоузнаваемой. Эти корковые нейроны делают огромный шаг вперед в процессе извлечения признаков, начатом в сетчатке. Данный конкретный признак, ориентированный край, создает упрощенное схематическое изображение объекта, передача которого в разы более экономичный процесс, чем передача полного изображения. Это можно сравнить с векторным и растровым способом представления изображений в компьютерной графике. Изображения в векторном формате имеют гораздо меньший объем и передаются гораздо быстрее, чем растровые, в которых записывается и передается каждый пиксель. Растровые изображения – самый полный, но и очень неэффективный способ передачи графической информации.

Второй тип клеток, который Хьюбел и Визель назвали «сложными» клетками, также реагировал на линии и края определенной ориентации, но был менее требовательным к их местоположению. Эти клетки возбуждались, когда край имел правильную ориентацию, но был привязан к более широкой зоне. На рисунке, как и в предыдущем примере, вертикальные штрихи обозначают отдельные спайки, генерируемые клеткой, когда в ее рецептивное поле попадает линия с правильным наклоном. Когда же линия имеет другой угол наклона, клетка молчит.
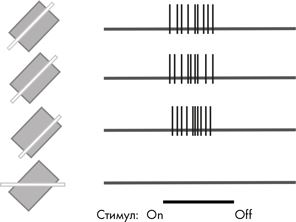
Итак, обобщим: простая клетка возбуждается светлым (или темным) краем определенной ориентации, который находится в конкретном месте поля обзора. Сложная клетка также чувствительна к краю определенной ориентации, но с некоторой степенью свободы: она возбуждается, если такой край появляется где угодно в пределах довольно обширного рецептивного поля, а не только в узкой области.
Вышесказанное важно, потому что эти клетки, по сути, занимаются вычленением такого абстрактного признака, как линейность, до некоторой степени не привязанного к конкретному визуальному стимулу. Хотя их рецептивные поля все равно ограничены, эти клетки выявляют линейность в относительно обширной области, а не в конкретном месте. Это возвращает нас к проблеме, упомянутой в начале книги: к нашей способности распознавать букву А независимо от того, где ее изображение падает на центральную область нашей сетчатки. В 1960-х гг. был разработан последовательный иерархический метод распознавания более сложных объектов, основанный на переходе от обычных неориентационных клеток к простым и далее к сложным избирательным в отношении ориентации клеткам. Эта модель работала не очень хорошо, однако сам механизм, отличающий сложные клетки от простых, сыграл ключевую роль в изобретении важного типа компьютерного зрения (подробнее обо всем этом мы поговорим чуть позже).
Дальше наш путь лежит в почти неизведанные края – в кору головного мозга. Наше понимание коры, говоря откровенно, находится на детсадовском уровне. Пока у нас есть лишь отдельные островки знаний – знаний об отдельных корковых областях, функции которых нам хотя бы приблизительно известны. К счастью, постепенно эти островки начинают соединяться в единый ландшафт – очень грубую, но все же целостную картину того, как организована система зрительного восприятия в головном мозге.
7 | Что дальше: лоскутное одеяло зрительной коры
Есть известные неизвестные вещи – те, о которых мы знаем, что мы их не знаем. Но есть также неведомые неизвестные вещи – такие, о которых мы не знаем, что мы их не знаем… Последние, как правило, представляют наибольшую трудность.
Неврологи и нейробиологи считают, что кора головного мозга является средоточием всех тех способностей, которые делают нас людьми, – таких как мышление, речь и чувства. Неудивительно, что зрительная кора в последние десятилетия так интересует исследователей, занимающихся изучением зрения. Важный прорыв был сделан в конце 1990-х гг. вследствие усовершенствования безболезненных способов регистрации нейрональной активности у обезьян, которые находились в сознании и выполняли определенные действия. Это дало нам возможность соотнести конкретные визуальные стимулы с активностью нейронов в конкретных зрительных областях головного мозга.
До сих пор, говоря о «коре», я имел в виду первичную зрительную кору – основную мишень аксонов, выходящих из латерального коленчатого тела. Первичная зрительная кора, сокращенно обозначаемая V1 (от англ.
Итак, оказалось, что зрительные области в головном мозге похожи на лоскутное одеяло. Разные области по-разному реагируют на визуальные объекты и соединены между собой малопонятной нам схемой связей. Поскольку основной массив исследований был сосредоточен на обезьянах и их зрительная кора на сегодняшний день изучена лучше всего, далее я буду говорить в основном об обезьянах, предварительно напомнив вам о том, что, судя по анатомическим и другим параметрам, человеческое зрение не слишком сильно отличается от обезьяньего.
На рисунке на следующей странице показан головной мозг, на котором отмечены отдельные зрительные области. Каждая из этих областей выполняет свою задачу в системе обработки визуальной информации, такую как распознавание объектов, обнаружение движения и т. д.
Первичная зрительная кора (V1) – основной пункт назначения зрительной информации, поступающей из латерального коленчатого тела. Зоны V2, V3 и V4 постепенно уходят все дальше в мозг; их можно рассматривать как более-менее последовательные звенья в цепи обработки зрительной информации.
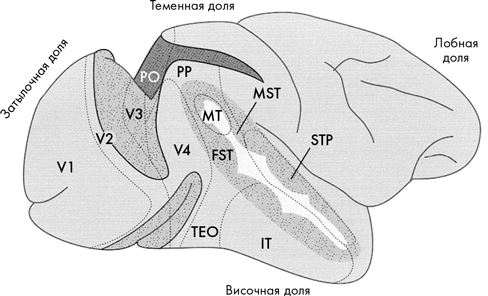
Помимо них существует множество других зрительных областей, обозначенных на схеме аббревиатурами, некоторые из которых я расшифрую по ходу повествования[22].
Некоторые исследователи считают, что в зрении так или иначе задействовано около 30 % всего объема мозга. Но что же делают все эти вычислительные центры? Эта система так сложна и многообразна, что моему мозгу не хватает когнитивной мощи охватить всю ее сразу. Поэтому я начну с двух наиболее изученных корковых областей: средней височной области (почти в центре рисунка), которая связана с восприятием движения и замечательной группы пятен в нижней височной коре, чьи клетки специализируются на распознании лиц.
Множество умнейших людей корпело над изучением работы нейронов в средней височной коре. Помните мой рассказ о том, как постдоки регистрировали электрическую активность клеток в латеральном коленчатом теле? Здесь происходит все то же самое: вы подводите микроэлектрод вплотную к нейрону, воздействуете на сетчатку различными видами визуальных стимулов и, отслеживая активность нейрона, пытаетесь выяснить, на какой именно стимул он реагирует. Так вот, исследования показали, что нейроны средней височной коры обладают свойствами, которые мы не обнаруживали ранее.
Прежде всего рецептивные поля средневисочных нейронов оказались намного больше, чем у нейронов сетчатки и даже у нейронов первичной зрительной коры. В зависимости от техники измерений их рецептивные поля в четыре – десять раз превышают в размере поля нейронов первичной коры V1. Это означает, что средневисочные нейроны больше нельзя рассматривать как зрительные пиксели; они занимаются более абстрактными вещами.
Ученые выяснили, что эти клетки делают кое-что удивительное. Большинство средневисочных нейронов избирательны в отношении направления: как и нейроны сетчатки, они сообщают остальному мозгу, что что-то движется в определенном направлении. Но из-за большого размера своих рецептивных полей они (в отличие от нейронов сетчатки) не локализуют движущийся объект – тот может находиться где угодно в довольно обширной области видимого мира, чтобы вызвать усиленное возбуждение. Однако эти нейроны обладают другими весьма полезными свойствами.
В пределах своих рецептивных полей средневисочные нейроны относительно неразборчивы. Они реагируют на движение не только одного большого объекта, но и множества маленьких точечных объектов. По причинам, в которые я не буду здесь вдаваться, нейроны сетчатки плохо реагируют на такой тип раздражителя. Можно подразнить средневисочный нейрон, показав ему облако точек, часть которых движется в одном направлении, часть – в обратном (обычно для генерации таких сложных стимулов мы используем специальную компьютерную программу). Так вот, средневисочный нейрон реагирует только в том случае, если решает, что точек, движущихся в предпочитаемом им направлении, больше, чем точек, движущихся в противоположную сторону.
Итак, эта клетка сообщает остальной зрительной системе, что что-то движется в определенном направлении, не уточняя, что именно движется и в какой именно точке пространства. Что еще любопытнее, некоторые средневисочные нейроны реагируют на движение частично невидимых объектов, например таких как барберполы – «цирюльничьи столбы», которыми цирюльники в прошлые века обозначали свои заведения. Барберпол – это вертикальный цилиндр, вращающийся вокруг горизонтальной оси. Но из-за того, что на цилиндр нанесен узор из косых линий, при вращении создается впечатление, будто линии движутся вверх (или вниз, в зависимости от направления вращения). Разумеется, это зрительная иллюзия; на самом деле каждая точка на поверхности цилиндра описывает абсолютно горизонтальный круг вокруг оси вращения. Специализированные лабораторные эксперименты с барберполом показали, что именно средневисочные нейроны обнаруживают это несуществующее движение полос вверх.