Относительно координат третьей оси конечности задаются различия между ладонью и ее тыльной стороной, а также определяется направление, в котором будет сгибаться локтевой сустав. И на этот раз почка конечности использует источник сигнальных молекул, расположенный на одной из ее сторон.[199] Сигнальной молекулой на этот раз служит белок из семейства WNT (WNT7a), который синтезируется только с той стороны конечности, которая в дальнейшем станет тыльной стороной ладони. WNT7a подавляет формирование признаков, характерных для ладони: у мутантных животных, лишенных функционального гена WNT7a, структуры, характерные для ладони, развиваются с двух сторон конечности.[200]
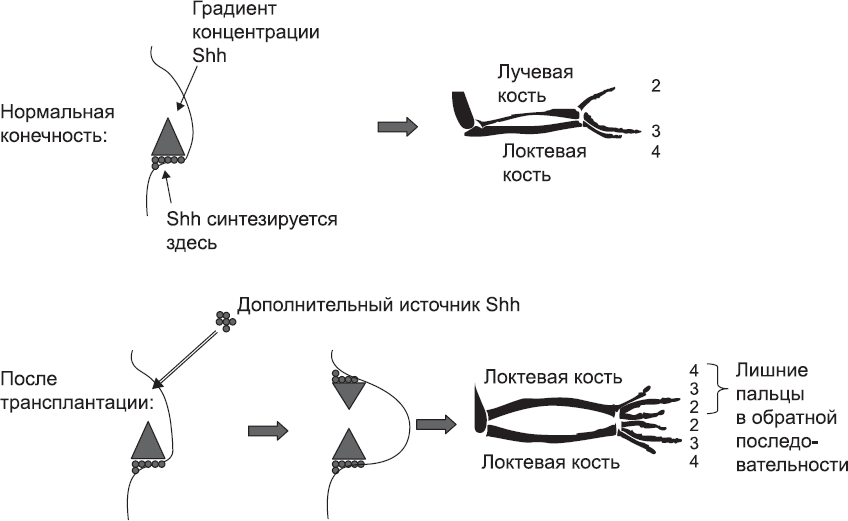
Рис. 55. В ходе нормального развития почки передней конечности цыпленка образуется крыло с тремя пальцами, гомологичными первому, второму и третьему пальцу человека. Если поместить на противоположной стороне почки крыла второй источник Shh, обычный комплект пальцев дублируется в зеркальном отражении, а обе кости предплечья развиваются в локтевые кости
Итак, складывается впечатление, что конечность контролирует свою разметку при помощи градиентов концентрации сигнальных молекул. Градиенты расположены под прямым углом друг к другу, охватывая все три пространственных измерения. Геометрически эти оси независимы, но с точки зрения биохимической активности они сплетены тесным и еще непонятным нам образом.[201] Экспериментальное вмешательство в одну из осей нарушает, по крайней мере частично, и формирование структур относительно двух других осей. Сигнал WNT7a, определяющий положение структур относительно оси «ладонь – тыльная сторона руки», необходим для производства нормального количества белка Shh со стороны мизинца. Хотя бы небольшое количество белка Shh нужно для поддержания нормального уровня FGF. В свою очередь, Shh и FGF необходимы для поддержания нормального уровня WNT7a. Наших знаний о развитии конечностей недостаточно, чтобы понять, зачем нужна такая взаимозависимость. Возможно, она помогает поддерживать правильное соотношение сигналов, поступающих из разных источников.
По мере роста конечностей возникает потребность в эффективной кровеносной системе, обеспечивающей транспорт кислорода и питательных веществ к размножающимся клеткам. Новые сосуды образуются в ответ на сигналы от ткани точно так же, как и в других системах (см. главу 9). Проблема в том, что конечности удлиняются очень быстро, и кровеносная система должна формироваться так же быстро, чтобы от них не отставать. В противном случае клеткам конечности будет не хватать питательных веществ, и их размножение замедлится, что приведет к образованию маленьких, укороченных, аномальных конечностей. Печальной иллюстрацией этого служат последствия одной медицинской ошибки, пустившей под откос жизнь примерно десяти тысяч людей в 1958–1961 гг.
Причиной трагедии стало благое намерение смягчить или даже полностью устранить утреннюю тошноту. Это состояние, нередкое на ранних сроках беременности, может быть крайне изнурительным. Очень эффективным в борьбе с утренней тошнотой оказался один препарат, уже использовавшийся в качестве успокоительного и противовоспалительного средства, и его стали активно назначать беременным женщинам. Это был талидомид. В 1958 г. еще не знали (на самом деле выяснилось это только в начале этого века), что молекула, образующаяся в результате распада талидомида, ингибирует рост новых кровеносных сосудов.[202] Это достаточно сильный эффект, и если будущая мать принимает талидомид в то время, когда быстро растущие конечности больше всего нуждаются в новых кровеносных сосудах, образование сосудов начнет отставать, и рост конечности замедлится. В крайнем случае это может привести к полному отсутствию конечностей или к образованию маленьких кистей или стоп непосредственно на туловище. Не совсем понятно, почему кисти рук и стопы продолжают развиваться, когда остальная часть конечности уже не растет, но если недостаток питательных веществ заставляет клетку задержаться в зоне прогресса, это явление хорошо согласуется с временной моделью формирования конечности.
На выявление связи между внезапным всплеском рождения младенцев с деформированными и укороченными конечностями и талидомидом ушло два года. К 1961 г. связь была доказана, и талидомид перестали прописывать от утренней тошноты. Следует заметить, что его продолжают использовать как лекарство от некоторых других болезней, включая проказу, и один-два врожденных дефекта в год (в среднем по миру) связаны с его применением в этих целях. Талидомид вернулся и на Запад, так как он эффективен при некоторых глазных и раковых заболеваниях, причем именно за счет способности подавлять рост кровеносных сосудов. Однако следует крайне внимательно следить за тем, чтобы препарат не назначался женщинам, которые потенциально могут забеременеть.
Глава 12
X или Y? Вот в чем вопрос
Продолжение человеческого рода – великое чудо и таинство. Если бы Господь спросил моего мнения по этому вопросу, я посоветовал бы ему и дальше вылеплять новых людей из глины.
В предыдущих главах речь шла о том, что пережил каждый из нас без исключения, о том, что происходило с эмбрионом до возникновения памяти, индивидуальности и даже половых различий. В течение первых семи недель развития определить пол будущего ребенка по каким бы то ни было внешним признакам невозможно, и только хромосомный анализ может приоткрыть завесу этой тайны. Мужские и женские тела формируются из одного и того же материала; возникновение частей тела, характерных для одного из двух полов, не связано с усилением каких-то изначальных анатомических различий, поскольку таких различий нет. Это процесс, в ходе которого некоторые ткани эмбриона выбирают один из двух возможных путей развития – мужской или женский. У млекопитающих первыми выбирают этот путь половые железы (гонады), которые затем «сообщают» о своем решении другим частям организма.
Яичники женщин и семенники (яички) мужчин состоят из большого количества различных типов клеток, которые для простоты можно разделить на две группы. Во-первых, это клетки зародышевой линии – яйцеклетки или сперматозоиды и клетки, которые дают им начало. Только клетки зародышевой линии могут передать генетический материал новому человеку, поэтому в некотором смысле в них заключается суть репродуктивной системы. А если, как это делают некоторые биологи, считать, что жизнь – это передача генов из поколения в поколение, то в этих клетках заключается суть человеческой жизни. Как заметил Сэмюэль Батлер, «курица – это приспособление, с помощью которого одно яйцо производит другое». Вторая группа клеток половых желез известна под общим названием соматических клеток. Это название связано с тем, что эти клетки относятся к уже существующему телу («соме»), и ни они, ни их потомки не могут дать начало организму нового поколения. К соматическим клеткам относятся клетки, продуцирующие половые гормоны, клетки, обеспечивающие защиту и поддержку клеток зародышевой линии в процессе развития в сперматозоиды и яйцеклетки, а также многие другие клетки, которые поддерживают целостность гонад, обеспечивают их кровоснабжение и иннервацию, а также выполняют множество других рутинных функций.
Соматические клетки гонад и клетки зародышевой линии возникают в разных частях эмбриона. Соматические клетки гонад образуются по обе стороны от средней линии в верхней части туловища – примерно на уровне нижних долей легких, если мы представим в виде эмбриона самих себя. Такое положение гонад у эмбриона, резко контрастирующее с их локализацией у взрослых людей, особенно у мужчин, может показаться странным, но в свете эволюции оно вполне логично. Люди, как и другие млекопитающие, рептилии, птицы и земноводные, произошли от рыб. Даже у взрослых рыб, особенно у «примитивных» видов, гонады расположены в передней части тела. Там они могут взаимодействовать с формирующимися кровеносными сосудами и другими тканями, что жизненно важно для создания функциональной репродуктивной системы. Эти взаимодействия необходимы и в человеческом организме, а поскольку ткани, вовлеченные в процесс формирования гонад, участвуют и в формировании аорты (глава 9), они не могут сместиться без серьезнейших последствий для всей кровеносной системы. Поэтому и место формирования половых желез никуда передвинуться не может.
Клетки зародышевой линии (первичные половые клетки) впервые выявляются перед самым началом гаструляции, когда в эпибласте на заднем конце первичной полоски обосабливается группа из примерно пятидесяти клеток (глава 4). Вероятно, эти клетки вовлекаются в гаструляцию на очень ранних этапах, а затем они движутся вместе с мезодермой, которая выходит за пределы тела эмбриона.[203],[204] Пока в процессе гаструляции образуются туловище и голова (см. главу 4), клетки зародышевой линии находятся в верхней части желточного мешка (рис. 56). Там они и остаются, пока продолжается формирование тела эмбриона и его частей и структур: нервной трубки, сомитов, кровеносной системы и т. д. Когда основной план строения уже намечен, зародышевые клетки возвращаются обратно, в тело эмбриона, как за счет морфогенетических движений, связанных с формированием кишечника, так и за счет активной миграции. Внешняя поверхность развивающейся кишечной трубки служит им столбовой дорогой, по которой легко добраться от задней части тела до области развивающихся гонад, которые секретируют особый набор сигнальных молекул. Привлеченные этими молекулами, зародышевые клетки заползают в гонады. По ходу дела зародышевые клетки пролиферируют, и вместо пятидесяти пустившихся в путь клеток к гонадам прибывает пятитысячная армия (впоследствии клеток зародышевой линии станет еще больше).
Хотя именно клетки зародышевой линии дадут начало сперматозоидам и яйцеклеткам, пол эмбриона определяют не они, а соматические клетки половых желез. Процесс «принятия решения» о выборе пола основан на взаимодействии нескольких белков. Их названия неудобочитаемы, но необходимы для описания механизма определения пола, так что заранее прошу прощения.
На этом этапе развития конкретная группа соматических клеток половой железы начинает производить белок WT1. Он выполняет множество разных функций в клетках, но к самым важным относится связывание определенных последовательностей ДНК и включение вместе с другими ДНК-связывающими белками определенных генов. Большинство активируемых WT1 генов находятся на хромосомах, которые есть у каждого человеческого эмбриона, но один из них – ген SRY – находится на Y-хромосоме,[205] которая есть только примерно у половины эмбрионов (почему это так, я объясню чуть позже). Если у данного эмбриона Y-хромосома есть, WT1 запустит образование белка SRY. Если же она отсутствует, то не будет и белка SRY. Это различие определяет весь дальнейший ход событий.
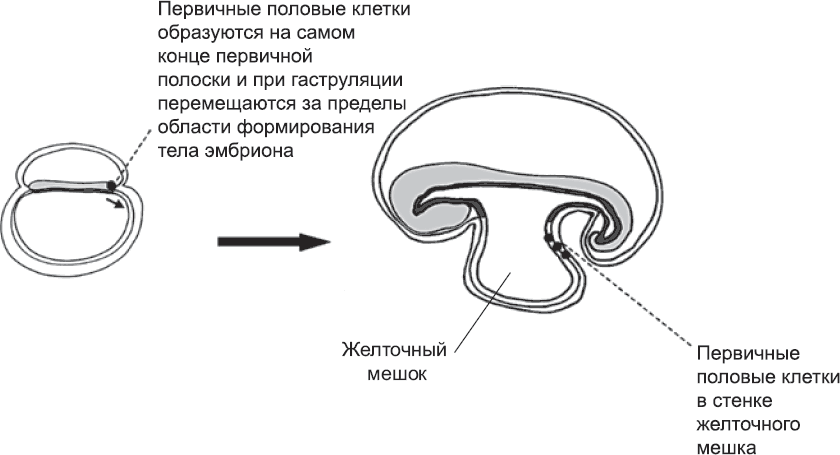
Рис. 56. Ранние клетки зародышевой линии (первичные половые клетки) образуются в задней части первичной полоски и перемещаются в желточный мешок, где ждут, пока идет процесс формирования тела эмбриона
Давайте сначала рассмотрим эмбрион, в котором может образоваться белок SRY. Он, как и WT1, связывается с определенными участками ДНК, но с другими, чем WT1. SRY активирует новые гены, которые, не будь этого белка, остались бы неактивными. Один из этих генов кодирует белок SOX9, а он в свою очередь активирует экспрессию очередной «порции» генов. Таким образом, различия в характере экспрессии генов между клеткой с Y-хромосомой и клеткой без нее растут как снежный ком.[206] Отметим, что ген SOX9 находится не на Y-хромосоме, а на семнадцатой хромосоме, которая есть у каждого эмбриона.
Прежде всего белок SOX9 обеспечивает дальнейшее производство самого себя. Это важно, так как эмбрион должен принять четкое и бесповоротное решение по поводу своего пола, в противном случае может образоваться организм, сочетающий мужские и женские черты. Экспрессия SOX9 активирует сигнальный путь, основанный на белках FGF,[207] а этот путь, в свою очередь, обеспечивает дальнейший синтез SOX9 даже в отсутствие SRY.[208] Таким образом, петля SOX9 – FGF может поддерживать сама себя, и как только она активирована, Рубикон перейден: эмбрион встал на мужской путь развития (рис. 57).
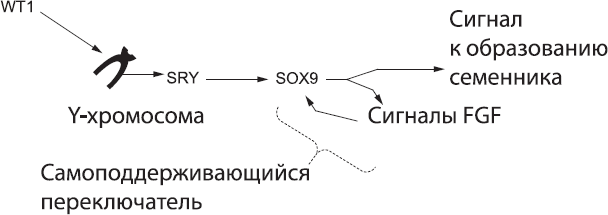
Рис. 57. Биохимический каскад «самцовости», который активирует переключатель SOX9 – FGF
Важность петли SOX9 – FGF была подтверждена в ряде экспериментов над генетически модифицированными мышами. Эмбрионы мышей, в клетках гонад которых Sox9[209] полностью отсутствовал, формировали признаки, характерные для самок, несмотря на присутствие Sry, так как Sry не может влиять на организм без работоспособного Sox9.[210] Такой же эффект наблюдался после удаления сигнальной системы Fgf. Напротив, мыши, в гонадах которых экспериментально активировали экспрессию Sox9 независимо от присутствия Sry, развивались в самцов, даже если у них не было Y-хромосомы.[211]