Когда аксоны встречают на своем пути другую клетку, они образуют особый межклеточный контакт – синапс, – через который передается их сигнал. Иногда синапсы основаны на непосредственном электрическом контакте с клеткой. Однако чаще встречаются так называемые химические синапсы: аксон выпускает в небольшую щель между собой и другой клеткой нейромедиатор – мелкую молекулу, стимулирующую рецепторы принимающей клетки, а они, в свою очередь, стимулируют электрическую и/или биохимическую активность в клетке. Так осуществляется передача сигнала. Разные типы нейронов используют разные нейромедиаторы. Именно работу нейромедиаторов имитируют или ингибируют медицинские препараты (а также наркотики), воздействующие на работу мозга за счет избирательного повышения или понижения активности некоторых его систем.
Основная задача развивающейся нервной системы – правильно «протянуть» все эти «провода». Нейроны должны надежно соединяться друг с другом и, когда это необходимо, с органами чувств (глаза, уши, нос, кожные рецепторы и т. д.) или с другими органами (мышцы, кровеносные сосуды, железы и т. д.). Существенную часть этой важной работы выполняют специальные структуры, расположенные на концах растущих аксонов, – конусы роста.
Конусы роста (рис. 65) состоят в основном из белков, которые обеспечивают миграцию клеток (см. главу 8).[226] У них есть ведущий край, где микрофиламенты организованы в сеть, способную продвигать его вперед. По мере сборки микрофиламенты сети удлиняются и толкают мембрану. Иногда продвижение сопровождается образованием длинных и тонких филоподий, способных вытягиваться далеко за пределы конуса роста, а затем снова втягиваться. Двигательные белки, расположенные в глубине конуса роста (например, миозин), взаимодействуют с микрофиламентами, организуя их в сократимые пучки, которые тянут лидирующий край назад. Если бы ничто не препятствовало этому, передний край конуса роста мог просто сокращаться, и никакого движения вперед не происходило бы. Однако конус роста также несет комплексы белков, прикрепляющихся к определенным участкам поверхности, по которой продвигается конус.[227] Эти белковые комплексы надежно заякоривают систему микрофиламентов, обеспечивая точку отталкивания для ведущего края. Благодаря этим комплексам не миозин тянет ведущий край в обратном направлении, а наоборот, центральная часть конуса роста подтягивается к ведущему краю. Таким образом, конус роста, а значит и аксон в целом продвигаются вперед.
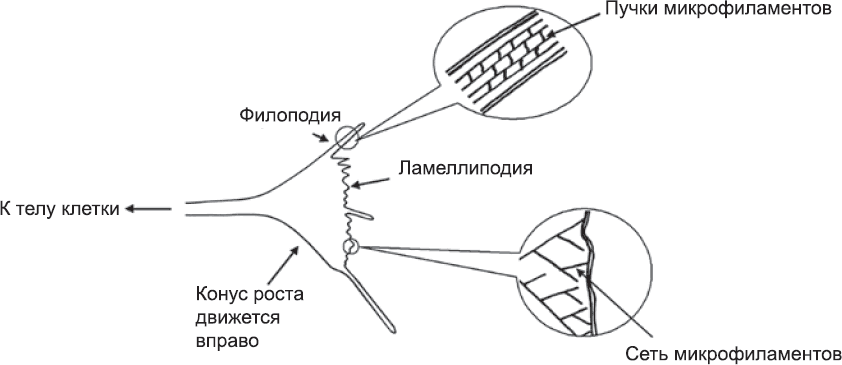
Рис. 65. Микрофиламенты конуса роста организованы в два типа структур: ламеллиподию (лидирующий край), которая поддерживается разветвленной сетью отдельных микрофиламентов, и филоподии, которые удлиняются, выталкиваясь за счет роста пучков микрофиламентов
Описанный выше механизм предполагает наличие тесной связи между способностью конусов роста крепиться к поверхности и их способностью продвигаться вперед. Это особенно важно, когда конус роста встречает на своем пути разные поверхности, а именно это и происходит как в экспериментальных условиях, так и в живых эмбрионах. Если поверхность под конусом роста более «липкая» с одной стороны конуса, чем с другой, он сместится в сторону более «липкой» поверхности. Именно по этой поверхности будет наиболее эффективно проталкиваться вперед новый лидирующий край. В результате конус роста будет двигаться в этом направлении.
Разные типы нейронов синтезируют разные типы белков адгезионного комплекса. Более того, один и тот же нейрон на разных этапах своего существования может производить разные наборы белков адгезионного комплекса. Каждый получившийся из этих наборов белков адгезионный комплекс избирательно прикрепляется к определенной молекуле на поверхности, по которой движется нейрон (если эта молекула там есть). Таким образом, если перед разными нейронами открывается один и тот же выбор поверхностей, они могут начать расти в разных направлениях.
Различия в адгезивных свойствах поверхностей – не единственная навигационная подсказка, которой могут воспользоваться конусы роста. Некоторые другие сигналы воздействуют на молекулярные механизмы, обеспечивающие сборку лидирующего края конуса роста.[228] Если на различные участки конуса роста действуют разные концентрации внешних сигнальных молекул, баланс между удлинением и сокращением на этих участках будет разным, и конус роста будет двигаться в направлении сигналов, активирующих удлинение, и удаляться от сигналов, активирующих сокращение.[229] Иногда разница в концентрации сигнальных молекул в разных областях эмбриона настолько велика, что конус роста реагирует по типу «все или ничего». Тогда конусы роста определенного типа вообще не достигают определенного участка. В других случаях различия в концентрации не столь резки, и конусы роста реагируют на них более сложным образом, что позволяет плавно подводить их к нужной цели. Бывает и так, что конусы роста доходят до нужного участка эмбриона по градиенту концентрации.
Пример реакции по типу «все или ничего» мы находим в системе, которая определяет, будут ли аксоны пересекать среднюю линию спинного мозга. Строгий контроль в этой системе очень важен, например, для того чтобы человек мог совершать асимметричные движения руками – скажем, для того чтобы, удерживая блюдце в левой руке, правой поднести к губам чашку чая. При движении руки с чашкой происходит произвольное сокращение бицепса (и других мышц). Насколько нам известно, молекулы, которые синтезируются в бицепсе левой и в бицепсе правой руки, совершенно одинаковы. Конусы роста двигательных нейронов в спинном мозге, предназначенные для управления бицепсами, сами по себе не могут уловить разницу между правой и левой рукой. Если бы конусы роста могли свободно пересекать центральную ось спинного мозга перед тем, как покинуть его и отправиться к мышцам, многие нейроны, которые должны контролировать правую руку, контролировали бы и левую, и наоборот. Тогда руки могли бы совершать только одинаковые движения. Этого необходимо избежать, а значит, двигательные нейроны, отходящие от правой стороны спинного мозга, должны идти только к правой руке. То же самое относится и к органам чувств. Мы можем понять, слева или справа от нас находится источник звука, потому что мозг соответствующим образом связан с сенсорной системой левого и правого уха. Организм может обнаружить и исправить какие-то случайные ошибки при помощи компенсаторных механизмов (о них пойдет речь в главе 15), но эти механизмы работают только в том случае, если большая часть «проводки» была изначально проложена правильно.
Пересечет ли конус роста среднюю линию (предположим, например, что речь идет о конусе роста аксона вставочного интернейрона, который передает сигналы от одного нейрона спинного мозга к другому), зависит главным образом от его реакции на контакт с клетками вентральной пластинки (напомню, что эта структура имеет форму полоски, которая проходит по вентральной поверхности нервной трубки; см. главу 5). На поверхности клеток вентральной пластинки есть белок под названием SLIT. Его распознает рецептор ROBO, имеющийся в конусах роста некоторых нейронов. При связывании рецептора ROBO с белком SLIT внутри конуса роста запускаются сигнальные пути, которые блокируют продвижение лидирующего края клетки, а также приводят к его быстрому сокращению.[230] Итак, если какая-то часть лидирующего края конуса роста, несущего рецептор ROBO, вошла в контакт с клеткой, экспрессирующей SLIT, то она сокращается, и продвижение аксона в этом направлении блокируется. Аксон будет расти в том направлении, куда его ведет та часть лидирующего края, которая не контактировала с несущими белок SLIT клетками средней линии, и поэтому не сможет ее пересечь. Если же конус роста лишен рецептора ROBO, он абсолютно невосприимчив к белку SLIT и может безнаказанно пересечь среднюю линию (рис. 66). Набор белков, экспрессирующихся в конусе роста, может меняться на протяжении его жизни. Аксоны, которые должны пересечь среднюю линию, могут послужить тут отличным примером. Пока они движутся к средней линии, у них есть рецепторы, которые регистрируют «притягивающие» сигналы клеток средней линии, а рецепторов ROBO практически нет (более того, у них синтезируются белки, которые в любом случае подавляют работу рецепторов ROBO). Поэтому такие аксоны без труда пересекают срединную линию. В процессе пересечения, однако, их конусы роста подвергаются воздействию высоких уровней концентрации белка Sonic Hedghog, который синтезируют клетки вентральной пластинки (глава 7). Под действием этого белка конусы роста после небольшой задержки, достаточной для пересечения средней линии, приобретают чувствительность к ROBO.[231] Это означает, что средняя линия теряет для них привлекательность, и они не пытаются снова пересечь ее, а движутся дальше к местам назначения.[232] (Аббревиатура ROBO происходит от названия мутации плодовых мушек
Если клеткам нужно образовывать связи только со «своей» стороной центральной нервной системы, они образуют аксоны, которые не должны пересекать вентральную пластинку. Конусы роста этих аксонов несут рецепторы ROBO и не могут пересечь «негостеприимную» зону вентральной пластинки. Некоторые из этих нейронов чувствительны как к «привлекающим» сигналам вентральной пластинки, так и к «отпугивающим» белкам SLIT. Столкнувшись с этим противоречивым влиянием, они остаются как можно ближе к сигналу-аттрактанту, но все же не настолько близко, чтобы непосредственно столкнуться с сигналом-репеллентом.[233] Подобно мотылькам, которые танцуют вокруг пламени, но не сгорают, они держатся на постоянном расстоянии от вентральной пластинки и растут параллельно ей вдоль спинного мозга. Такие аксоны связывают между собой разные уровни тела, например мозг с иннервирующими бицепсы моторными нейронами, располагающимися на уровне рук.
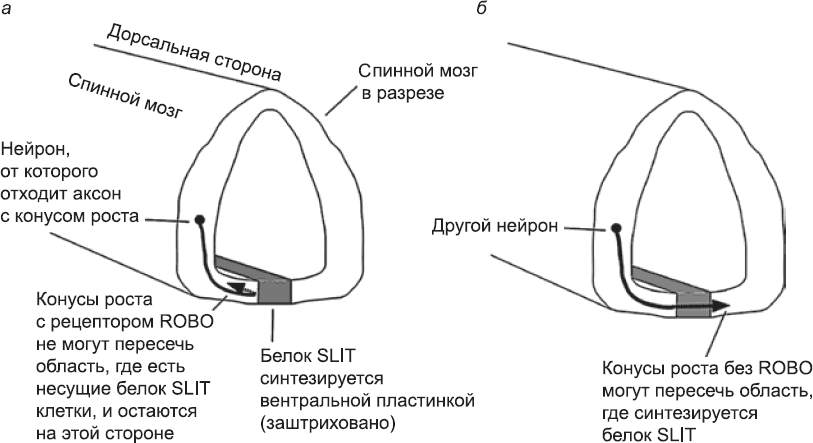
Рис. 66. Вентральная пластинка спинного мозга вырабатывает сигнальную молекулу SLIT. Она «отпугивает» конусы роста, несущие активный рецептор ROBO (
Примером реакции, основанной на оценке относительных концентраций сигнальных молекул-репеллентов и плавно подводящих нейроны к цели, может служить установление связи между глазами и мозгом. В глазу взрослого человека изображение проецируется на сетчатку – изогнутый «экран», выстилающий глазное яблоко сзади. Сетчатка содержит светочувствительные клетки – напряжение на их мембранах меняется в зависимости от яркости падающего на них света. Светочувствительные клетки соединены с залегающими в непосредственной близости нейронами, которые выполняют локальную обработку сигналов перед передачей их в мозг. У каждого из этих нейронов есть аксон, идущий прямо в мозг. Многие из них идут в ту часть мозга, которая у млекопитающих называется верхним двухолмием.[234] Аксоны ганглиозных клеток сетчатки идут параллельно друг другу в виде толстого «кабеля» – зрительного нерва, но на подходе к верхнему двухолмию они расходятся и подключаются к нему очень необычным образом. Место в верхнем двухолмии, к которому подключается аксон, точно соответствует положению на сетчатке ганглиозной клетки, от которой он отходит. По сути, структура верхнего двухолмия точно повторяет расположение ганглиозных клеток на сетчатке, поэтому на нем отображается – в виде сигналов электрической активности – точная копия оптического изображения сетчатки.
В нанесении «карты» сетчатки на верхнее двухолмие при эмбриональном развитии участвуют несколько механизмов. Сначала наносятся общие контуры, а затем «карта» постепенно прорабатывается и становится очень точной. Один мощный механизм основан на взаимодействии молекул-репеллентов на поверхности клеток верхнего двухолмия с рецепторами конуса роста, которые при обнаружении таких молекул вызывают локальное разрушение лидирующего края.[235] Производство молекул-репеллентов клетками двухолмия неравномерно. Их очень много в той его части, которая должна в конечном итоге соединиться с аксонами носовой (то есть ближайшей к носу) стороны глаза. По мере удаления от этой части клетки двухолмия синтезируют все меньше таких молекул, а в той части, которая должна в конечном итоге соединиться с аксонами ближайшей к уху (височной)[236] стороны сетчатки, их совсем мало. Неравномерно и производство рецепторов конусами роста ганглиозных клеток сетчатки: конусы роста аксонов клеток носовой стороны образуют очень мало рецепторов, но по мере удаления от нее их количество неуклонно растет, и конусы роста на височной стороне глаза производят гораздо больше рецепторов. Конусы роста аксонов клеток на височной стороне сетчатки будут активно избегать клеток верхнего двухолмия, которые содержат большое количество белка-репеллента. Поэтому они будут двигаться к краям верхнего двухолмия, где этого белка мало. Конусы роста аксонов клеток, расположенных ближе к середине глаза, тоже несут некоторое количество молекул рецептора, а значит, будут отклоняться от той части верхнего двухолмия, клетки которого синтезируют много молекул-репеллентов. Однако верхнее двухолмие имеет ограниченный размер, и аксоны, идущие от клеток середины сетчатки, не могут соревноваться с аксонами, идущими от височной стороны сетчатки (где образуется больше всего рецепторов), для подключения к той части, где молекул-репеллентов очень мало. В результате им приходится «довольствоваться» областями, где таких молекул не слишком много. Конусы роста на носовой стороне сетчатки имеют так мало рецепторов, что способны «вытерпеть» даже область верхнего двухолмия с очень высоким содержанием молекул-репеллентов. Благодаря такой «конкуренции» за возможность уйти подальше от молекул-репеллентов – а она тем сильнее, чем конусы роста чувствительнее, – конусы роста располагаются в двухолмии в том же порядке, в каком они покинули сетчатку (рис. 67).
Описанный выше механизм позволяет упорядочить конусы роста в соответствии с их положением на горизонтальной (нос – ухо) оси глаза. Похожая система, использующая другие пары рецепторов и молекул-репеллентов, организует конусы роста относительно вертикальной оси (лоб – щека), так что на верхнем двухолмии оказывается точная двумерная «карта» сетчатки. Вполне вероятно, что вдоль обеих осей действуют и другие, пока неизученные сигнальные системы, основанные не только на репеллентах, но и на аттрактантах, и они осуществляют тонкую доработку карты, нанесенной с помощью описанного выше механизма. Мы находимся в самом начале пути к пониманию этих процессов развития.
Механизм «подключения» конусов роста аксонов зрительного нерва к нужному месту в головном мозге, о котором я только что говорил, основан на локальных взаимодействиях. Однако не следует забывать, что, прежде чем конусы роста оказались в районе верхнего двухолмия, они уже проделали долгий путь от сетчатки к верхнему двухолмию. Этот путь тоже очень сложен и основан на множестве разнообразных «отпугивающих» и «привлекающих» сигналов.[237]
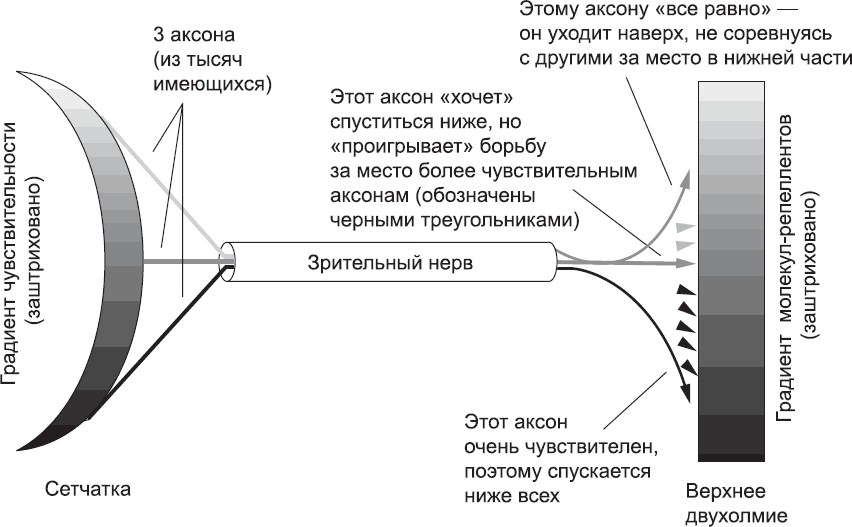
Рис. 67. Схема, поясняющая механизм, благодаря которому «карта» сетчатки наносится на верхнее двухолмие (из тысяч аксонов показаны лишь три). Аксоны клеток на височной стороне сетчатки (нижняя часть схемы) несут большое количество рецепторов и активно избегают молекул-репеллентов в верхнем двухолмии. Поскольку эти молекулы распределяются по градиенту, аксоны избегают всего двухолмия, кроме той части, где этих молекул меньше всего (на схеме внизу). Аксоны, отходящие от средней части сетчатки, тоже избегают высоких концентраций молекул-репеллентов, но не столь активно. Таким образом, они остаются посередине верхнего двухолмия (они бы и рады спуститься ниже, но им не дают этого сделать аксоны с височной стороны). Аксоны клеток, лежащих на носовой стороне сетчатки, образуют мало рецепторов и почти не реагируют на молекулы-репелленты. Поэтому они подключаются к той области верхнего двухолмия, где этих молекул много (никакие другие аксоны на эти места не претендуют). Следует подчеркнуть, что на рисунке представлена очень упрощенная схема этого процесса, не отражающая ни сложный путь зрительного нерва, ни очертания верхнего двухолмия
Первая навигационная задача, с которой сталкиваются конуса роста на пути от сетчатки глаза к мозгу, – найти точку в задней части глаза, из которой должен выходить зрительный нерв. Это достигается за счет того, что у конусов роста есть рецепторы, чувствительные к молекулам-репеллентам, которые распространяются от края сетчатки, и рецепторы к молекулам-аттрактантам, которые находятся ближе к центру сетчатки (откуда и будет выходить зрительный нерв) (рис. 68). Одной из таких молекул-аттрактантов является белок Sonic Hedghog. Если нарушить синтез этого белка в центре сетчатки эмбриона животного, то конусы роста аксонов ганглиозных клеток сетчатки не смогут обнаружить нужную точку и будут расти хаотично. Есть также данные, указывающие на существование градиента молекул-репеллентов между краем и центром сетчатки.
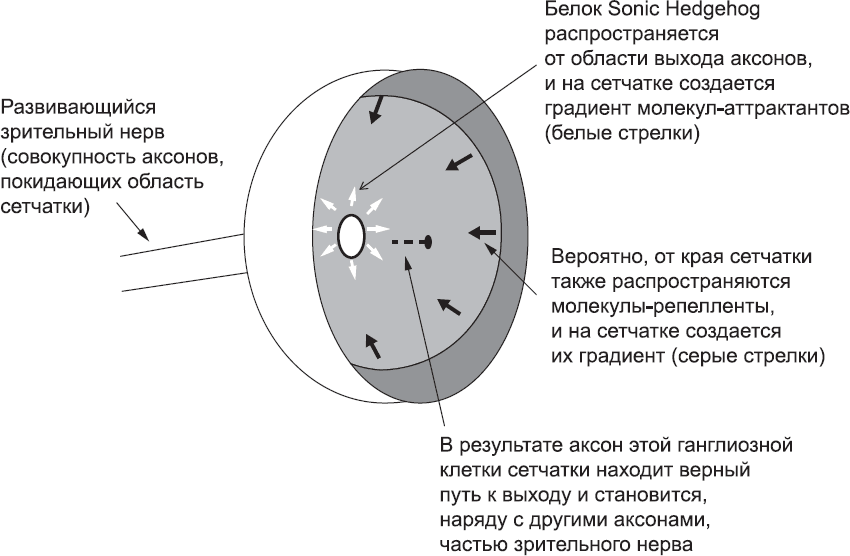
Рис. 68. Поиск аксоном пути в сетчатке, расположенной в задней части глаза и показанной на рисунке в виде чашеобразной структуры (примерно такую форму она и имеет). Аксоны ганглиозных клеток сетчатки находят путь к точке выхода, ведущей к зрительному нерву, пользуясь сигналами молекул-аттрактантов (таких, как Sonic Hedgehog), синтезируемых в центральной части сетчатки, а также, возможно, сигнальных молекул-репеллентов, синтезируемых по краям сетчатки
Первые конусы роста, покидающие область глаза, оказываются в узком «коридоре». Его стены образованы клетками, которые синтезируют еще один тип молекул-репеллентов.[238] По этому узкому коридору конусы роста могут двигаться только в одну сторону – в направлении центральной части развивающегося мозга. Когда тысячи конусов роста выходят из развивающейся сетчатки, тянущиеся за ними аксоны образуют мощный «кабель» – зрительный нерв.
Зрительные нервы от обоих глаз сходятся в одном и том же месте центральной части мозга и сталкиваются с необходимостью выбора дальнейшего маршрута. Некоторые из них пересекут срединную линию и направятся к верхнему двухолмию на противоположной стороне мозга, а другие направятся к верхнему двухолмию на своей стороне (рис. 69). Решающая, функциональная причина такого выбора маршрута связана с тем, как мы видим. У многих животных, особенно у тех, которые должны спасаться от хищников, глаза размещены по бокам головы.