Каждое мышечное волокно состоит из небольших структур, называемых мышечными фибриллами, или миофибриллами (
Зачастую эпимизий, перимизий и эндомизий простираются за пределы мясистой части мышцы, так называемого брюшка, формируя толстое «кабельное»[1] сухожилие или широкую, плоскую, слоистую сухожильную ткань, известную как апоневроз. Сухожилие и апоневроз формируют непрямые места «прикрепления» мышц к надкостнице или соединительной ткани других мышц. Более сложные мышцы могут иметь несколько мест прикрепления, например, четырехглавая мышца имеет четыре крепления. Поэтому, как правило, мышца перекрывает сустав и крепится по обе стороны кости при помощи сухожилий. Один конец мышцы остается в относительно зафиксированном или стабильном положении, в то время как другой ее конец двигается в результате мышечного сокращения.
Каждое мышечное волокно иннервируется единичным двигательным волокном, которое заканчивается недалеко от центра мышечного волокна. Единичное двигательное волокно и все мышечные волокна, которые оно задействует, являются двигательной единицей. Количество мышечных волокон, задействованных единичным двигательным волокном, зависит от движения, которое необходимо выполнить. Когда требуется точная, контролируемая степень подвижности, например движение глазом или пальцем, задействуется лишь несколько мышечных волокон; при необходимости выполнить более масштабное движение, например движение такими крупными мышцами, как большая ягодичная мышца, может быть задействовано несколько сотен мышечных волокон.
Отдельные мышечные волокна работают по принципу «все или ничего», когда стимуляция волокна приводит к полному его сокращению или к полному отсутствию такого сокращения – волокно не может сократиться чуть-чуть. Общее сокращение любой отдельно взятой мышцы предполагает сокращение определенного количества ее волокон в определенный момент времени, при этом остальные волокна находятся в расслабленном состоянии.
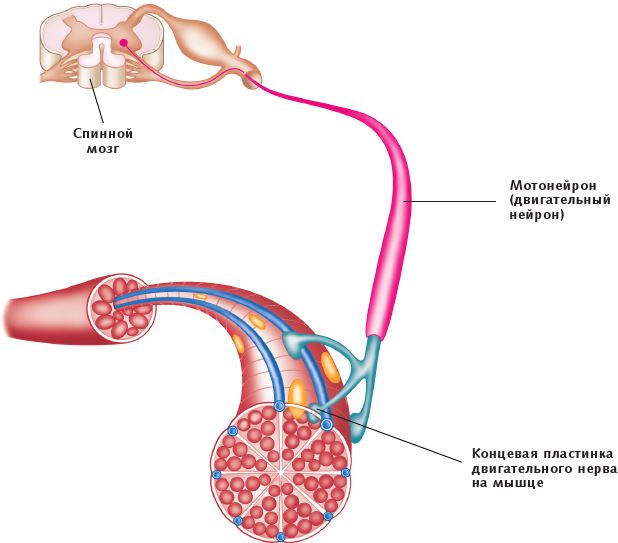
Рис. 1.5. Двигательная единица мышечного волокна скелетной мышцы
Физиология мышечного сокращения
Нервные импульсы приводят к сокращению мышечных волокон. Соединение между мышечным волокном и двигательным нервом известно как нейромышечное соединение, и именно здесь осуществляется взаимодействие между нервом и мышцей. Нервный импульс передается на нервные окончания, называемые синаптическим окончанием аксона, рядом с сарколеммой. В таких окончаниях содержатся тысячи пузырьков, наполненных нейромедиатором ацетилхолином (АХ). Когда нервный импульс достигает синаптического окончания аксона, сотни этих пузырьков высвобождают свой АХ, также и АХ открывает каналы, в которых происходит рассеивание ионов натрия (Na+). Потенциал покоя неактивного мышечного волокна составляет примерно –95 мВ. Инфлюкс ионов натрия уменьшает заряд, создавая потенциал концевой пластинки. Если потенциал концевой пластинки достигает порогового значения потенциала (примерно –50 мВ), ионы натрия попадают в поток, вследствие чего внутри волокна создается потенциал действия.
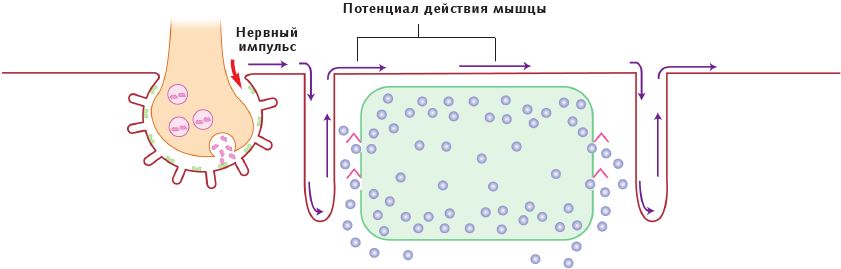
Рис. 1.6. Нервный импульс, представляющий собой триггер потенциала действия или сокращения мышцы
В мышечном волокне не происходит видимых перемен во время (и сразу после) потенциала действия. Этот период, называемый латентным, длится от 3 до 10 мс. Перед окончанием латентного периода фермент ацетилхолинэстераза (АХЭ) расщепляет АХ в нейромышечном соединении, натриевые каналы закрываются, и поле очищается в ожидании следующего нервного импульса. Потенциал покоя волокна восстанавливается путем оттока ионов калия из возбужденной клетки. Короткий период, требуемый для восстановления потенциала покоя, называется рефрактерным периодом.
Так каким же образом укорачивается мышечное волокно? Этот механизм можно лучше всего объяснить при помощи теории скользящих нитей (Huxley & Hanson, 1954), согласно которой мышечные волокна получают нервный импульс (см. выше), что приводит к выделению ионов кальция, сосредоточенных в саркоплазматическом ретикулуме (СР). Чтобы мышцы работали эффективно, нужна энергия, которая создается в результате распада аденозинтрифосфата (АТФ). Такая энергия позволяет ионам кальция связываться с филаментами актина и миозина для формирования магнитной связи, в результате чего волокна укорачиваются, вызывая сокращение мышц. Мышечное действие продолжается вплоть до истощения запасов кальция, после чего кальций начинает возвращаться в СР, где он будет храниться до следующего нервного импульса.
Мышечные рефлексы
В скелетных мышцах содержатся специальные сенсорные единицы, восприимчивые к удлинению (укорачиванию) мышцы. Такие сенсорные единицы называются мышечным веретеном и нервно-сухожильным веретеном (сухожильным органом Гольджи), они важны для обнаружения изменений в длине мышцы, реагирования на такие изменения и для их регулирования.
Мышечные веретена состоят из спиральных нитей, которые называются интрафузальными мышечными волокнами, а также нервных окончаний, расположенных внутри оболочки соединительной ткани для регулирования скорости удлинения мышцы. Если мышца удлиняется слишком быстро, сигналы, поступающие из интрафузальных мышечных волокон, уведомят об этом нервную систему через спинной мозг, чтобы нервный импульс был отправлен обратно, вызывая тем самым сокращение мышцы. Сигналы постоянно направляют в мышцу и из мышцы информацию, касающуюся положения и силы (проприорецепция).
Кроме того, когда мышца удлиняется и удерживается в таком положении, сократительная реакция будет сохраняться до тех пор, пока растянута мышца. Такой механизм известен как дуга разгибательного рефлекса. Мышечные веретена будут стимулироваться во время проведения растяжения мышцы.
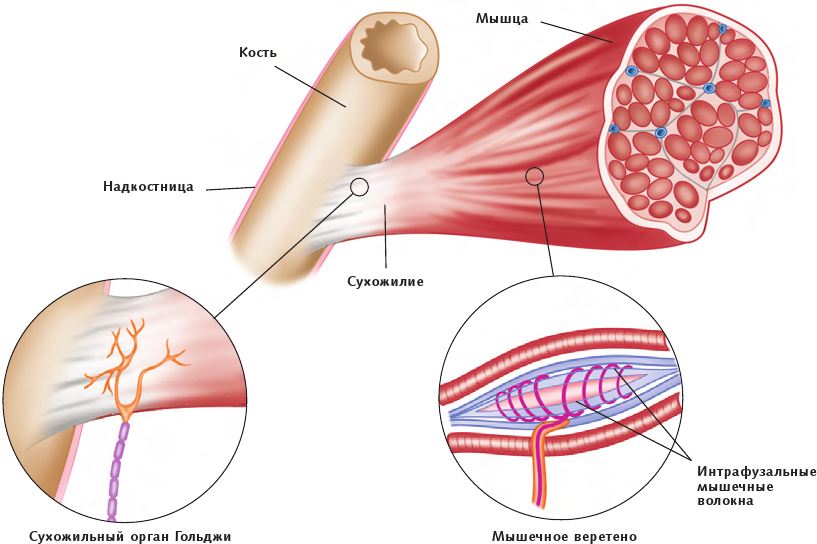
Рис. 1.7. Анатомия мышечного веретена и сухожильного органа Гольджи
Классический клинический пример разгибательного рефлекса – коленный рефлекс, который предполагает активацию рецептора растяжения сухожилия, что вызывает рефлекторное сокращение прикрепленной мышцы, то есть четырехглавой мышцы.
В то время как мышечные веретена контролируют длину мышцы, сухожильные органы Гольджи (СОГ) настолько чувствительны к напряжению в мышечно-сухожильном комплексе, что могут отреагировать на сокращение единичного мышечного волокна. СОГ по своей природе являются ингибирующими, выполняя защитную функцию и снижая риск травмы. При получении стимуляции СОГ ингибируют (тормозят) сокращающиеся мышцы (агонисты) и возбуждают мышцы-антагонисты.
Механика скелетно-мышечной системы
В большинстве случаев скоординированные движения предполагают прикрепление скелетной мышцы, которая остается в относительно стационарном состоянии с одной стороны и двигается с другой стороны места прикрепления. Проксимальное, стационарное прикрепление считается источником, а более дистальное, подвижное прикрепление считается вставкой. (В любом случае в настоящее время предпочтительнее употреблять именно выражение «место прикрепления» вместо «источник» и «вставка», поскольку мышцы устроены таким образом, что любой конец мышцы может двигаться или быть зафиксирован в зависимости от ситуации.)
В большинстве случаев движение требует задействования определенной мышечной силы, генерируемой мышцами-агонистами (или первичной движущей силой), которые в первую очередь отвечают за движение и обеспечивают большую часть силы, необходимой для осуществления движения. В движении также принимают участие мышцы-антагонисты, которые, удлиняясь, гарантируют движение, производимое первичной движущей силой, и выполняют защитную функцию. Кроме того, потребуется участие и мышц-синергистов (известных как стабилизаторы), помогающих первичной движущей силе и также порой участвующих в корректировке направления движения. Простым примером является сгибание локтевого сустава, требующее укорачивания плечевой мышцы и двуглавой мышцы плеча (первичная движущая сила) и расслабления трехглавой мышцы плеча (антагонист). Плечелучевая мышца выступает в качестве мышцы-синергиста, помогая плечевой и двуглавой мышцам плеча.