Чтобы подойти к теме гаструляции, можно было бы сначала рассмотреть строение примитивного организма, который образуется в результате данного процесса, а затем посмотреть, какой вклад в его образование вносит каждый этап гаструляции. Это, пожалуй, был бы самый рациональный и простой подход к описанию гаструляции, однако он может создать ложное впечатление о том, что клетки имеют какое-то представление об анатомии организма, в строительстве которого участвуют. На самом же деле развитие организма основано вовсе не на том, что клеткам известны законы развития (понять эти законы трудно даже нам, с нашим мозгом из миллионов клеток), а на том, что клетки автоматически откликаются на изменения окружающей их среды. Поэтому в этой главе мы, следуя логике развития, сначала рассмотрим поведение клеток развивающегося организма и только потом посмотрим, к чему оно приводит.
Прежде чем начать рассказ, хочу сделать важное предупреждение: изучать процесс гаструляции у человека крайне сложно, поэтому почти все, что описано в этой главе, основано на исследованиях животных. Существуют строгие правовые ограничения относительно выращивания человеческих эмбрионов в лабораторных условиях, и изучать эмбрионы на стадии гаструляции нельзя (почему – я расскажу чуть позже). Основная последовательность анатомических изменений человеческого эмбриона известна. Эти данные были получены в исследованиях немногочисленных (и очень ценных) эмбрионов, извлеченных при посмертном вскрытии или при удалении матки у женщин, которые чаще всего даже не знали о том, что беременны. Гаструляция начинается примерно через пятнадцать дней после оплодотворения, то есть примерно тогда, когда женщина ожидает начала менструации. Некоторым из этих эмбрионов более ста лет, но они до сих пор хранятся в музеях под особым присмотром, потому что новый материал такого рода поступает крайне редко. Исследования проводятся главным образом на мышах и курицах, а процесс гаструляции у этих животных значительно отличается от гаструляции у человека. Цыпленок развивается в яйце, а не в матке; эпибласт и гипобласт мыши имеют форму чаши, а не диска. Эти особенности могут играть существенную роль при развитии. Поэтому реконструировать механизмы гаструляции человека на основе данных, полученных при изучении модельных животных, рискованно – слишком мало у нас информации, и мы можем неверно интерпретировать какие-либо детали процесса.
Отправной точкой для гаструляции является эмбрион в том состоянии, в котором мы оставили его в конце главы 3. К этому времени уже сформировался ряд вспомогательных тканей, таких как плацента, а также два заполненных жидкостью внутренних пространства: амниотическая полость и желточный мешок. Между этими полостями лежат два диска, расположенные один над другим: гипобласт и эпибласт. Из гипобласта образуются дополнительные вспомогательные ткани, а из эпибласта – сам плод. Ни один из этих двух дисков не имеет каких-либо явных особенностей, которые позволяли бы отличить один его край от другого (см. рис. 9).
Первое изменение (судя по данным, полученным на животных) происходит в гипобласте. У клеток, расположенных в середине этого диска,[36] включаются новые гены, в том числе и ген ДНК-связывающего белка, который называется HEX.[37] Что является сигналом к этому изменению, пока неясно. Возможно, что все клетки гипобласта изначально готовы к нему, но большинство из них ингибируется сигнальным белком, который синтезируется тканями, окружающими гипобласт.[38] Достаточно далеко от источника этого ингибитора находятся только клетки в центре диска, поэтому они могут избежать его влияния и запустить экспрессию гена HEX. Этот механизм – только предположение, но активация гена HEX – факт. Клетки, в которых экспрессируется HEX, уходят из центра диска – они перемещаются, расталкивая соседние клетки, и собираются в одной точке на краю диска гипобласта[39],[40],[41] (рис. 10). Все еще непонятно, даже в случае мышей, что же особенного в этой точке диска, что клетки мигрируют именно туда. У «низших» животных расположение похожей «особой» точки обусловлено начальными условиями развития. В некоторых случаях положение этой точки определяется переходящей к зародышу асимметрией распределения молекул, заложенных в яйцеклетку материнским организмом. В других случаях в этой точке располагаются полярные тельца, «побочные продукты» клеточных делений, предшествующих формированию яйцеклетки. У некоторых организмов эту точку, по-видимому, маркирует место проникновения сперматозоида. Весьма вероятно, что у млекопитающих это тоже так, и есть указания на то, что эмбрионы мышей асимметричны уже на самых ранних стадиях развития.[42] Соответствующих данных по эмбрионам людей, разумеется, нет. Это очень досадно, потому что скопление в одном месте клеток, у которых экспрессируется HEX, приводит к исключительно важному результату – одно конкретное место на краю диска гипобласта становится не таким, как другие. Иными словами, это первый шаг в сторону от простой радиальной симметрии эмбриона (рис. 10).
Достигнув края диска гипобласта, группа клеток, экспрессирующих ген HEX, становится новой структурой ПВЭ.[43] Клетки ПВЭ начинают секретировать собственные сигнальные белки. Эти белки, способные распространяться на небольшие расстояния, вполне могут достигнуть эпибласта, лежащего непосредственно над гипобластом, явно в пределах дистанции распространения сигнала.[44] На этом этапе клетки эпибласта уже реагируют на сигналы, поступающие от вспомогательных тканей эмбриона. Эти сигналы подготавливают их к тому, чтобы создать структуры, характерные для задней части организма. Если бы больше ничего не происходило, весь эпибласт формировал бы структуры задней части, и это имело бы катастрофические последствия для эмбриона. Сигналы, поступающие от ПВЭ, уравновешивают эту тенденцию, запуская процессы создания тканей головы.[45] Если ПВЭ не будет синтезировать сигнальные белки, эмбрион не сможет правильно сформировать голову. Таким образом, положение ПВЭ в уникальной области на краю диска гипобласта диктует полярность вышележащего эпибласта: сторона, ближайшая к ПВЭ, сформирует голову, а удаленная от нее – заднюю часть тела эмбриона. Только небольшая часть эпибласта удалена от ПВЭ настолько, что вообще не получает от нее сигналов. По-видимому, это очень важно для локализации области, в которой происходят первые заметные изменения эпибласта. В этом месте клетки эпибласта переключаются на образование сигнального белка, который привлекает окружающие клетки – они мигрируют по направлению к источнику этого белка.[46] В результате такого перемещения клеток образуется важная структура со скромным названием – первичная полоска (см. рис. 10).
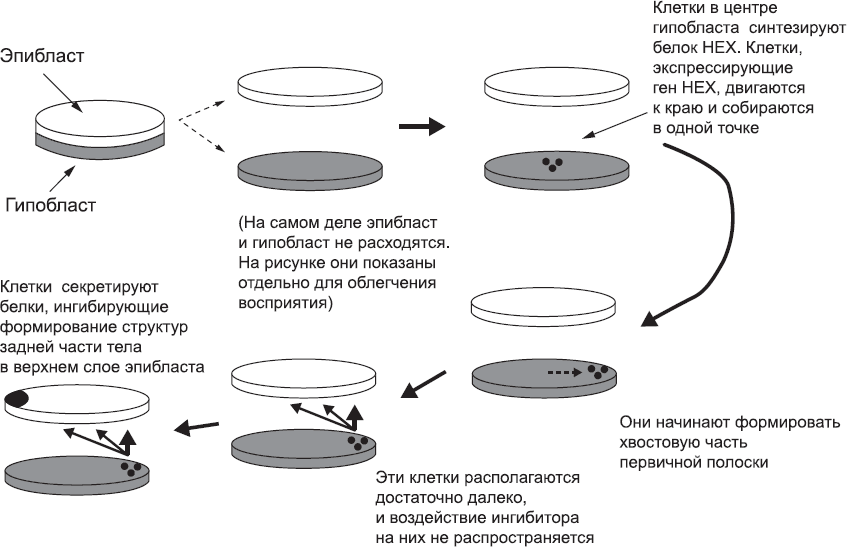
Рис. 10. Нарушение радиальной симметрии. Клетки в центре гипобласта «включают» ген HEX, движутся к краю и концентрируются в одном месте. Это нарушает радиальную симметрию гипобласта, а поскольку его клетки синтезируют сигнальные белки, которые «вмешиваются» в процессы, происходящие в эпибласте, его радиальная симметрия тоже нарушается
Особая роль ранней активности клеток ПВЭ гипобласта в определении места формирования первичной полоски была показана в экспериментах на куриных эмбрионах. После того как у них начинала нарушаться радиальная симметрия, гипобласт поворачивали относительно передне-задней оси эмбриона.[47] Эти манипуляции приводили к тому, что эпибласт формировал первичную полоску, подстраиваясь под новые координаты, а значит, именно гипобласт контролирует этот процесс.
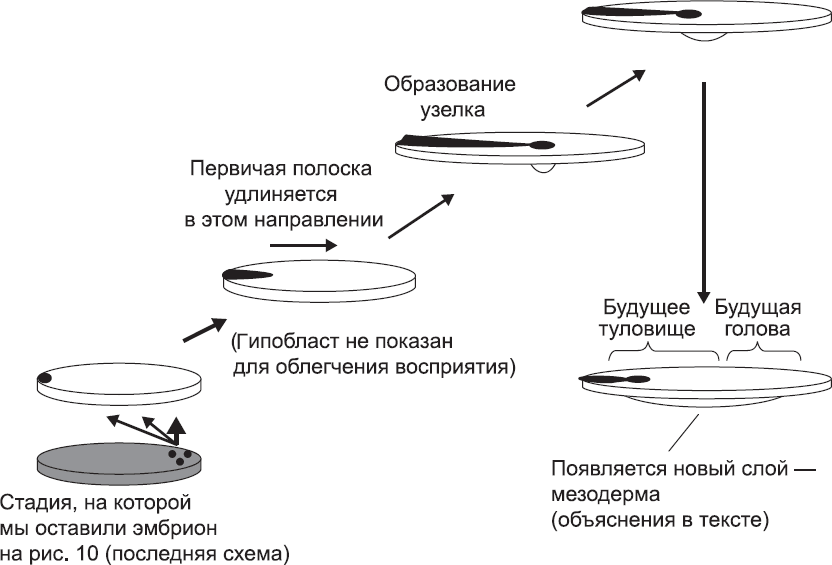
Рис. 11. Удлинение первичной полоски и образование узелка (что такое узелок, я объясню далее в этой главе)
Так как все больше и больше клеток эпибласта мигрирует в область первичной полоски, они начинают расталкивать друг друга и встраиваться между соседями. Это приводит к постепенному удлинению первичной полоски от края диска внутрь по его радиусу (рис. 11). Поскольку в эту миграцию вовлекается все больше клеток, на краях диска их становится меньше. В результате, по мере того как полоска удлиняется, диск в целом начинает сужаться. Он приобретает овальную форму, и радиальная симметрия эпибласта теряется (см. рис. 11). В длинной оси овала угадывается будущая ось тела, которая проходит от макушки до последнего позвонка (если бы речь шла о животных, можно было бы сказать «до кончика хвоста», но применительно к человеку это будет звучать немного странно). Та часть первичной полоски, которая образуется первой (она находится на краю диска), становится задней частью тела, а часть, которая образуется последней (примерно в середине диска, теперь имеющего овальную форму), становится головой. Таким образом, формирование нашего тела начинается с седалища, а создание головы откладывается «на потом», и это не единственный случай, когда эмбриология напоминает нам о том, что не стоит воспринимать себя слишком серьезно.
Роль ПВЭ в определении места образования первичной полоски имеет одно интересное следствие. Если клетки, экспрессирующие ген HEX, образуют не одно, а два скопления на краю диска, то сформируются две отдельные небольшие ПВЭ, а значит, два центра распространения сигналов. Тогда возможно образование двух первичных полосок. Это третий – и самый редкий – тип близнецов, который встречается менее чем в 1 % случаев двуплодной беременности.
Если образуются две независимые первичные полоски, то они дадут начало двум полностью независимым осям тела, а значит, двум организмам. Такие близнецы, возникшие из одного и того же эпибласта после образования амниотической полости, будут иметь общую амниотическую, а также хорионическую полость. Этим они отличаются от близнецов, описанных в главе 2 (у них нет общих полостей), и от близнецов, описанных в главе 3 (они имеют общую хорионическую полость, но амниотическая полость у каждого из них своя). (Именно по числу общих полостей акушеры могут понять, с каким случаем возникновения близнецов они имеют дело.) Возникновение близнецов за счет двух первичных полосок в одном и том же эпибласте чревато опасностями, так как между ними нет четкой разграничительной линии и существует серьезный риск, что они не разделятся полностью. В этом случае на свет появятся сиамские, или сросшиеся, близнецы. Как правило, они не просто «срослись» друг с другом, но и имеют общие части тела. Общими могут оказаться в том числе жизненно важные внутренние органы. В принципе, сросшиеся близнецы могут быть вполне здоровыми людьми, хотя, конечно, их жизнь простой не назовешь. Пожалуй, самые известные сиамские близнецы – это Чанг и Энг Банкер[48],[49] (1811–1874 гг.), много лет гастролировавшие с цирком Барнума. В те времена глазеть на уродов считалось нормальным, и бородатые женщины, карлики, великаны и аномально тучные люди в цирках выступали на потеху публики. Братья Банкер назвали свой «дуэт» в честь родной страны, и за феноменом сросшихся близнецов с тех пор закрепилось название «сиамские близнецы». Родись братья в наши дни, их, скорее всего, разделили бы, но во многих других случаях разделить сросшихся близнецов очень трудно или вообще невозможно. Иногда это возможно только ценой жизни одного из близнецов. Поэтому операции по разделению сросшихся близнецов, помимо чисто технических проблем, нередко осложняются серьезными этическими вопросами.
Еще более сложные ситуации возникают в тех случаях, когда эпибласт образует не две отдельные первичные полоски, которые дают начало двум осям тела, а одну «недоудвоившуюся» Y-образную первичную полоску. Это приводит к образованию двух голов и, возможно, двух шей при общем позвоночнике. Это явление называется «удвоением оси», но название явно неудачно, так как удваивается не вся ось (в этом случае проблем бы не было), а только ее часть. У людей это явление встречается редко – как правило, оно приводит к выкидышу или мертворожденному ребенку, а если младенец рождается живым, то скоро умирает. Заспиртованные экспонаты такого рода есть почти во всех старых коллекциях естественно-научных редкостей. Очень редко люди с этой аномалией все же выживают. Самый яркий пример – это сестры Абигейл и Бриттани Хенсел, которым сейчас около двадцати пяти лет. Голова и шея у каждой из сестер своя, а туловище общее. Все, для чего нужна голова, они могут делать совершенно независимо, например, каждая из сестер может читать независимо от другой. При ходьбе, игре на пианино и управлении автомобилем им приходится действовать вместе (надо полагать, чиновникам пришлось немало потрудиться над разработкой индивидуального экзамена по вождению). Несмотря на анатомические аномалии, сестры Хенсел абсолютно нормальные, умные и общительные молодые женщины. Удвоение оси более широко распространено у других животных, например рептилий и амфибий, которые выживают с этой аномалией гораздо чаще людей. Двуглавый крысиный полоз по имени Мы (We) прожил в Городском музее Сент-Луиса восемь лет и стал одной из его главных достопримечательностей. Известность ему принесли не только необычный внешний вид, но и «споры», время от времени возникавшие между его головами, каждая из которых имела свое мнение. Есть даже одна ископаемая находка взрослой двухглавой ящерицы.[50] Удвоение оси у амфибий можно индуцировать путем воздействия на сигнальные механизмы, эквивалентные сигнальным механизмам в эпибласте и гипобласте млекопитающих (собственно говоря, существование этих механизмов было впервые показано именно на лягушках). Например, в результате введения в эмбрион лягушки одного из белков, синтезируемого в ПВЭ млекопитающих и необходимого для формирования головы, получается лягушка с несколькими головами. Этот белок получил название Cerberus («Цербер») в честь многоголового пса, стража входа в Аид.
С точки зрения этики закономерность «одна первичная полоска – один человек» может быть истолкована как доказательство того, что ее образование – поворотный момент в развитии человека. Главным аргументом является то, что до формирования первичной полоски нельзя точно сказать, сколько людей получится из эмбриона, а значит, установить четкую связь между конкретным эмбрионом и конкретным человеком. Развивая эту мысль, можно прийти к выводу, что структура, которую нельзя приравнять к
Движения, благодаря которым клетки собираются вместе для формирования первичной полоски, не прекращаются и после ее создания. По мере поступления большего количества клеток центр полоски как бы проседает. Из полоски получается узкая бороздка, которая ближе к середине диска расширяется, образуя уплощенную область, называемую узелком. Клетки узелка продуцируют сигнальные белки, которые, во-первых, привлекают в этот регион дополнительные клетки, а во-вторых, заставляют их ослабить межклеточную адгезию и «включить» гены, способствующие их превращению в клетки других типов. Клетки, расположенные ближе к источнику сигнала (то есть к узелку), получают большую дозу сигнальных белков и реагируют сильнее. Ослабление системы адгезионных связей между клетками и активация у них миграционного поведения приводят к тому, что клетки свободно перемещаются в плоскости эпителиального диска и могут даже покинуть его.[51],[52],[53] Клетки проваливаются под поверхность диска эпибласта, переходя на его нижнюю сторону. Этот процесс распространяется по первичной полоске в направлении от головы к хвосту, так что более задние участки тела проходят его значительно позже, чем передние.[54] По мере того как ближайшие к центру бороздки клетки уходят из диска, их бывшие соседи сдвигаются ближе друг к другу и к центру бороздки так, чтобы заполнить свободное пространство, а затем, когда наступает их очередь, тоже проваливаются под диск, покидая эпибласт.
Первые клетки первичной полоски, которые уходят из диска на определенном уровне оси «голова – хвост», присоединяются к гипобласту: они расталкивают его клетки и встраиваются между ними, так что клеточный состав центральной области этого слоя полностью изменяется (рис. 12). Оказавшиеся в этой области клетки образуют слой, который называется эндодерма. (Это слово в переводе означает «внутренняя кожа»; название связано с тем, что в дальнейшем этот слой образует трубку кишки и трубки ассоциированных с ним органов – печени и поджелудочной железы.) Клетки, которые уходят из полоски немного позже, слабо связаны друг с другом и не образуют непрерывный слой. Они образуют мезодерму (в переводе – «средняя кожа»), которая лежит между эндодермой и внешним слоем. Клетки первичной полоски, которые не успевают «утонуть» и после завершения гаструляции остаются в эпибласте, формируют эктодерму (в переводе – «внешняя кожа»).[55] Таким образом, первичная полоска и узелок не только показывают нам ось «голова – хвост» у эмбрионов, но и превращают один слой клеток в три разных слоя: эктодерму, мезодерму и эндодерму (см. рис. 12). Это основные слои тела почти у всех животных.[56]
Для того чтобы уйти из диска, клетки должны сначала добраться до первичной полоски, и поэтому время их погружения тесно связано с исходным расстоянием до нее. Клетки, располагавшиеся ближе к средней линии диска, быстро проходят короткое расстояние и выселяются из полоски, тогда как клетки, располагавшиеся вблизи краев диска, начинают выселяться значительно позже, когда первые клетки уже давно погрузились под диск. Время и пространство так тесно сплелись в процессе гаструляции, что трудно определить, какой из этих факторов играет более важную роль в судьбе клеток. Определяется их судьба еще до начала миграции за счет ранних сигналов от ПВЭ или она становится известной только после того, как клетки выселяются из диска и оказываются внутри эмбриона? По-видимому, судьба некоторых клеток предопределена еще до начала движения.[57],[58],[59],[60],[61] Примером может служить обсуждавшееся выше программирование области, в которой будет формироваться голова, с помощью сигналов от ПВЭ, или программирование клеток, удаленных от ПВЭ, на формирование первичной полоски. Насколько четко определена на этих ранних этапах дальнейшая судьба других клеток, непонятно. Дело усложняется тем, что в некоторых случаях «запрограммированная» судьба клеток является всего лишь наиболее вероятным путем их развития. Если в эксперименте поместить их в другую среду и подвергнуть воздействию других сигналов (что вполне возможно и в естественных условиях, если во время гаструляции они затеряются среди других клеток), такие клетки могут «передумать» и пойти совсем по другому пути развития. Эта область эмбриологии все еще полна нерешенных вопросов, которые ждут своих исследователей.
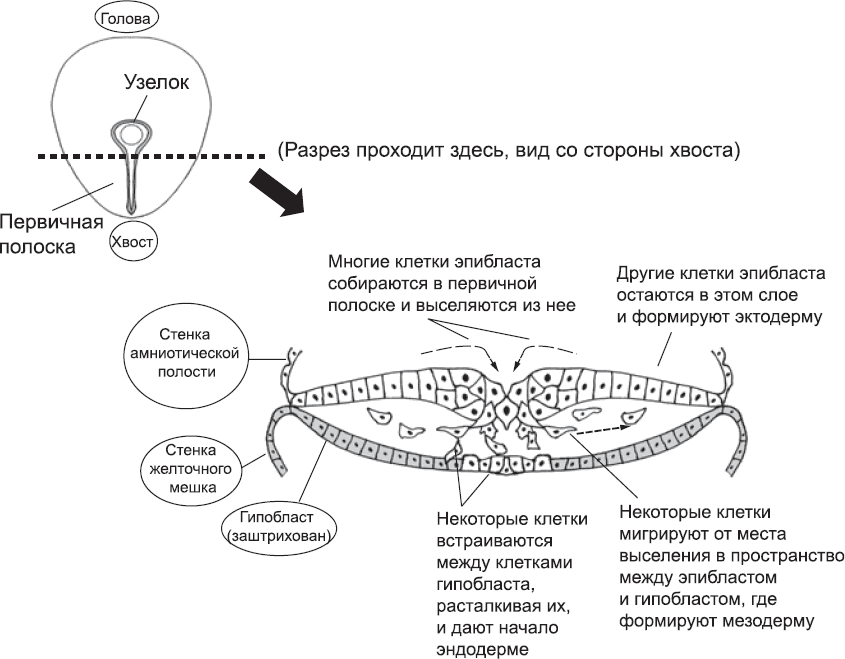
Рис. 12. Образование трех слоев тела – эктодермы, мезодермы и эндодермы – во время гаструляции. В верхней левой части изображен эпибласт эмбриона, у которого уже образовался узелок, как он виден со стороны амниотической полости. Гипобласт лежит под эпибластом и не виден. На основном рисунке изображена первичная полоска в разрезе (он обозначен пунктирной линией на рисунке в верхней левой части). Клетки эпибласта активно мигрируют в плоскости пласта, и траектории их движения сходятся в бороздке, получившейся из первичной полоски. Затем эти клетки выселяются из эпибласта и уходят в пространство между дисками. Часть из них (те, которые встраиваются между клетками гипобласта, расталкивая их от средней линии к краям) дадут начало эндодерме, а часть – мезодерме (новому слою клеток посередине между эпибластом и гипобластом). Клетки, не покинувшие эпибласт, формируют эктодерму
Почти сразу после образования эндодермы клетки, расположенные вдоль центральной линии диска гипобласта (то есть под первичной полоской), начинают двигаться вверх (рис. 13). Это клетки, которые выселялись из эпибласта непосредственно через узелок, с той его стороны, которая обращена к голове, а значит, это клетки, получившие самую высокую дозу сигнальных молекул узелка. Благодаря им эти клетки «запрограммированы» на то, чтобы покинуть эндодерму.[62],[63],[64] Сразу после этого они выстраиваются вдоль оси эмбриона и образуют плотный стержень – нотохорд.[65] Это одна из важнейших структур на ранних стадиях развития эмбриона. Чтобы понять, почему эта структура возникает так рано (а также то, почему она возникает вообще), нужно обратиться к эволюции животного мира.
Зоологи классифицируют животных, используя иерархическую систему. Основной единицей этой системы является вид (например,
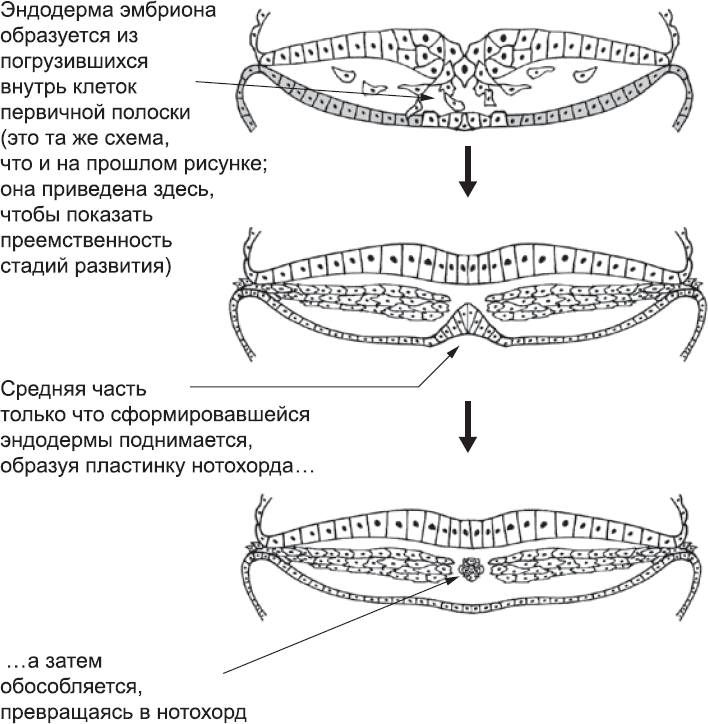
Рис. 13. Образование нотохорда из клеток, расположенных вдоль средней линии эндодермы (напомним, что эндодерма образовалась из погрузившихся клеток первичной полоски)